Introduction to cellular immune response
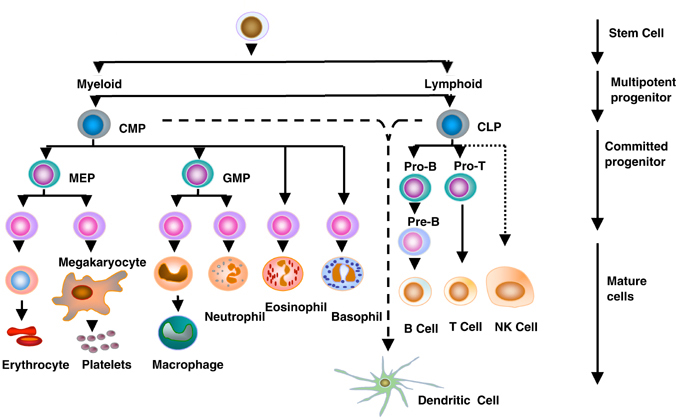
During embryonic development, the blood cell precursors originate mainly in the yolk sac and fetal liver. These cells differentiate into myeloid and lymphoid series. Lymphoid series leads to the development of B-lymphocytes, T-lymphocytes, and NK cells, and myeloid series leads to the formation of monocytes and macrophages, erythrocytes, neutrophils, basophils, eosinophils, megakaryocytes \platelets and dendritic cells. The normal ratio of T-lymphocytes: B-lymphocytes is T:B = 3:1. Lymphocytes, the cells competent to initiate immune responses, can be divided into two major groups: thymus-derived or T cells responsible for “cellular immunity” (e. g. delayed hypersensitivity reactions) and bursa (or bursa-equivalent) derived or B cells which produce immunoglobulin (antibody) molecules and are involved in “humoral immunity”. “Accessory” cells, such as monocytes (or macrophages), polymorphonuclear leukocytes and mast cells act in an auxiliary manner by facilitating antigen processing or presentation, or by liberating factors which modify the various manifestations of the immune response.
T-Lymphocytes
T lymphocytes arise from a pluripotent stem cell in the bone marrow. Unlike B-lymphocytes, which undergo continued maturation in bone marrow, T-lymphocytes differentiate in the thymus. B-lymphocytes have immunoglobulins on their cell surface through which they recognize antigens but T-lymphocytes are surface-negative for immunoglobulin and do not exhibit DNA rearrangements in immunoglobulin genes. Rather, T-lymphocytes express a distinct membrane receptor for the antigen that recognizes antigen in conjunction with membrane glycoproteins encoded in the major histocompatibility complex. T lymphocytes play a major role in the initiation and regulation of immune responses and are key elements of cell-mediated immune responses against viruses, intracellular bacteria, and tumors. T-lymphocytes express different surface membrane antigens at various stages of development and/or cell activation. These surface markers have been useful in the identification of phenotypic and functional diversity among T-lymphocytes. These markers previously were identified by means of monoclonal antibodies, which recognize specific antigenic determinants (termed epitopes) within a given surface membrane protein. So, a leukocyte surface marker, which is reactive toward a group (or cluster) of monoclonal antibodies, is identified according to a cluster of differentiation (CD) number (e.g. CD2, CD8). Certain CD markers are expressed by virtually all peripheral blood T-lymphocytes. This is the case with respect to CD2, a 50-kilodalton glycoprotein through which T-lymphocytes form rosettes with sheep red blood cells.
Other CD markers are useful in segregating T-lymphocytes into a number of distinct subpopulations. Thus, approximately 60 percent of peripheral blood T-lymphocytes express CD4, a glycoprotein expressed on T-cells whose activation is dependent on the recognition of antigen in conjunction with class II major histocompatibility complex (MHC) molecules. Approximately 30 percent of peripheral blood T-lymphocytes express CD8; a membrane protein expressed by T-cells whose activation is dependent on the recognition of antigen in conjunction with class I MHC molecules. The majority of circulating T-lymphocytes express either …… Contents available in the book………… Contents available in the book…… Contents available in the book…… Contents available in the book………
T-lymphocyte ontogeny
T-cell development occurs in the thymus. T-cell precursors, however, arrive at the thymus from the bone marrow. The stages of T-cell development are identified by the expression of specific cell surface markers, such as TCR (T Cell Receptor), CD3 (which serves as the signal transduction component of TCR), and CD4/CD8. In the cortex of thymus within the cortical epithelium, T-cells progenitors differentiate under the influence of thymic hormones (Thymosin and Thymopoietin) into T-cell subpopulation. All T-cells have CD-3 receptor on their surface in association with Antigen receptors. CD-3 is a complex of 5 trans-membranous proteins which takes information from outside to the inside of the cell. Signal transduction is by Zeta-chain (transmembranous protein) which has tyrosine kinase activity.CD-4 has single trans-membranous polypeptide whereas CD-8 has 2 transmembrane polypeptides. These may signal by tyrosine kinase.
T-lymphocytes undergo differentiation in the thymus, irrespective of whether they express the CD4 or CD8 phenotype. The process of T lymphocyte maturation begins with the migration of T-cell precursors from the bone marrow to the cortical regions of the thymus. It is thought that these cells are attracted to the thymus by chemical signals provided by thymic epithelial cells. The earliest recognizable thymocyte committed to the T-cell lineage is the pro-T-cell. These cells bear CD2 (the sheep erythrocyte receptor) but do not express the T-cell antigen receptor. Pro-T-cells are also surface-negative for CD4 and CD8 (Double Negative or DN cells).
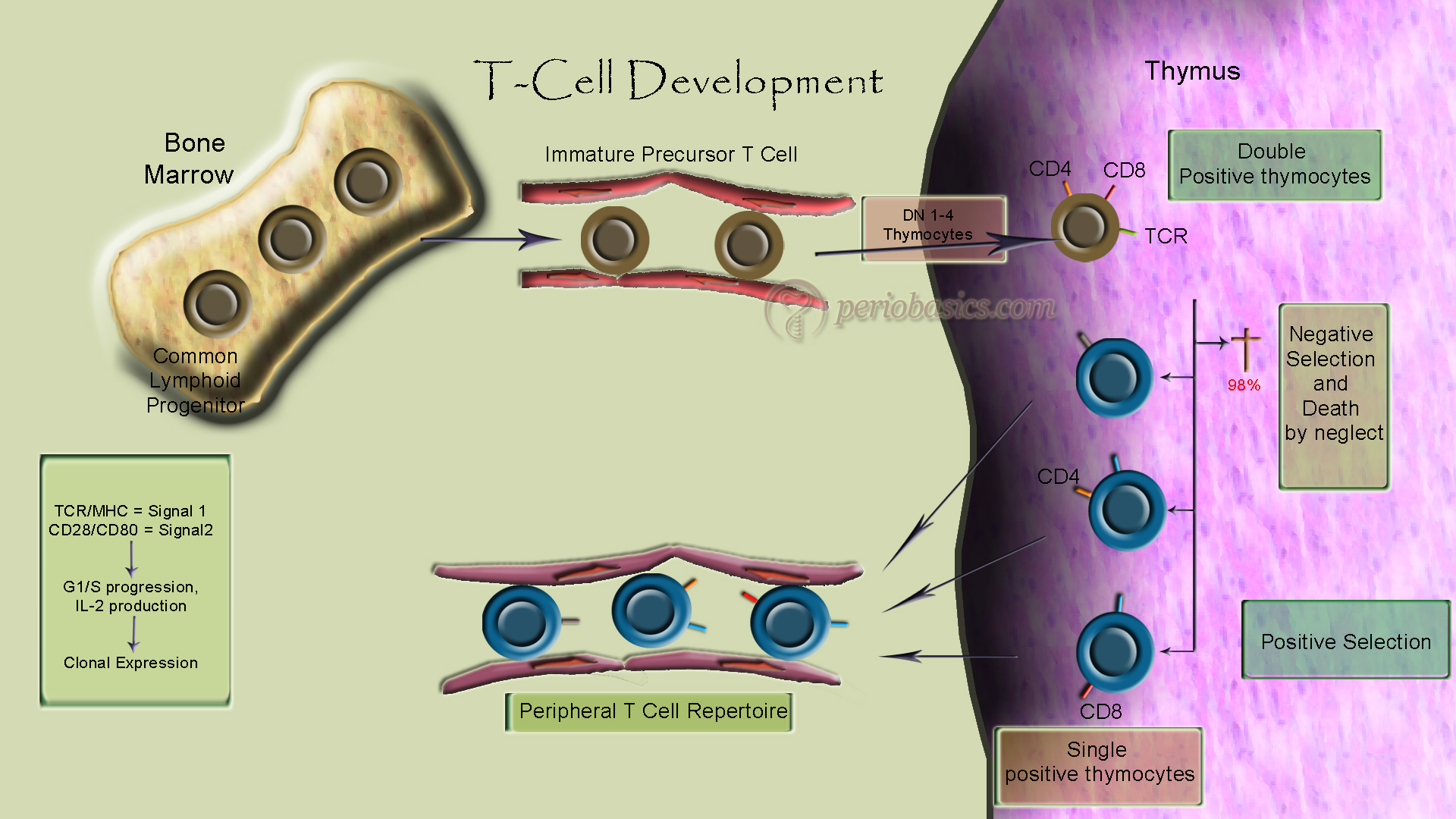
During the next phase of development, maturing cortical thymocytes proceed along one of two alternative pathways. In one instance, the thymocytes begin to rearrange DNA segments encoding for the variable and constant regions of an alternative form of the T-cell antigen receptor (designated λ / δ TCR) and express this receptor in conjunction with a tightly associated complex of five membrane glycoproteins that form the CD3 complex. Most cells expressing the λ / δ TCR fail to express either CD4 or CD8 during further development and release into the peripheral circulation as “double-negative” T-lymphocytes. The precise function of these λ / δ TCR–expressing cells has not been defined. The majority of pro-T-cells follow a different pathway of differentiation. This pre-T-cells co-express both CD4 and CD8 ( Double positive or DP cells), but do not yet express an antigen receptor. Subsequently, these cells undergo DNA rearrangements of genes encoding the constant and variable regions of the α / β TCR, which is expressed by the majority of mature T-lymphocytes in the periphery. At this stage, the thymocytes are CD4+, CD8 +, and express the α / β TCR in conjunction with the CD3 complex. During the first stages of development, which occur in the thymic medulla, two important events take place called thymic education.
Thymic education
It is an important step in the maturation of T-cells. First, the cells progressively lose either CD4 or CD8. Secondly, these cells are “educated” by thymic epithelial cells to learn to differentiate between “self” and “nonself”-MHC gene products. Those cells, which are auto-reactive toward self-MHC molecules, are eliminated by a process of clonal deletion, an important mechanism of self-tolerance. As a consequence of this selection process, only about 10 percent of the immature T-cells which enter the thymus eventually reach the peripheral circulation.
Positive selection
Since TCR’s recognize antigen only in the context of MHC’s, T-cells must be tuned to recognize host MHC first. During positive selection double-positive T-cells that can recognize self MHC’s are selected for proliferation, and those T cells that do not recognize self MHC die via Apoptosis. The positive selection also assures that the right TCR selection will go with the appropriate CD4 or CD8. For example, TCR’s specific for MHC II need to retain CD4 and lose CD8. If the reverse occurs, they will die via apoptosis. The same is true for the T cells that are specific for MHC I, which need to retain CD8 and lose CD4
Negative selection
At this point, those T-cells that are strongly activated by self MHC plus self-peptides need to be eliminated in the thymus. If they escape this elimination, they may subsequently react against self-antigens, and cause autoimmune disease.
In summary, Positive selection selects for those T cells that react with MHC: self-antigen. Negative selection eliminates those that react strongly with MHC: self-antigen. Thus, successful T cell differentiation selects for MHC restricted TCR’s with low affinity for self-antigens. Cells that fall outside this range primarily die via apoptosis. The rationale here is that a T cell that binds weakly to self MHC/self Antigen will not be activated but will be activated by a stronger binding to self MHC/ foreign Antigen complex.
Mature T-cells that survive this selection process leave the thymic medulla through the walls of postcapillary venules. After circulating for a time, these T-lymphocytes distribute among various …… Contents available in the book………… Contents available in the book…… Contents available in the book…… Contents available in the book………
Naïve Th cell
Naïve Th cell is a T-cell that has differentiated in the bone marrow and successfully undergone the positive and negative selection processes in the thymus. A naïve T-cell is considered to be mature and unlike activated T-cells or memory T-cells it has not encountered its cognate antigen within the periphery. Upon activation, naïve Th cells become Th0, since they have both Th1 and Th2 characteristics, with further stimulation Th0 cells deviate either towards Th1 or Th2. Th1 and Th2 cells are classified based on the pattern of cytokines that they secrete. If the Th cell secretes mainly IL2 and INF-γ, it is termed a Th1 cell. If the T cell secretes mainly IL4, IL10, and IL13 it is termed a Th2 cell.
Natural killer (NK)-cells
Natural killer (NK) cells are granular lymphocytes that do not pass through the thymus. They don’t have CD-4 or CD-8 proteins and an antigen receptor. They kill virus-infected and tumor cells without the requirement of antigen presentation or MHC proteins. NK-cells are activated by IL -2. Natural killer (NK) cells are lymphocytes of the innate immune system that are involved in early defenses against both allogeneic (nonself) cells and autologous cells undergoing various forms of stress, such as infection with viruses, bacteria, or parasites or malignant transformation. Although NK cells do not express classical antigen receptors of the immunoglobulin gene family, such as the antibodies produced by B cells or the T cell receptor expressed by T cells, they are equipped with various receptors whose engagement allows them to discriminate between target and nontarget cells. Activating receptors bind ligands on the target cell surface and trigger NK cell activation and target cell lysis. However Inhibitory receptors recognize MHC class I molecules (HLA) and inhibit killing by NK cells by overruling the actions of the activating receptors. This inhibitory signal is lost when the target cells do not express MHC class I and perhaps also in cells infected with a virus, which might inhibit MHC class I expression or alter its conformation. The mechanism of NK-cell killing is the same as that used by the cytotoxic T-cells generated in an adaptive immune response; cytotoxic granules are released onto the surface of the bound target cell, and the effector proteins they contain penetrate the cell membrane and induce programmed cell death.
Dendritic cells
Dendritic cells (DCs) were originally identified by Steinman and his colleagues in 1972. They are potent antigen-presenting cells (APCs) that possess the ability to stimulate naïve T cells. They comprise a system of leukocytes widely distributed in all tissues, especially in those that provide an environmental interface. DCs are derived from bone marrow progenitors and circulate in the blood as immature precursors prior to migration into peripheral tissues. Within different tissues, DCs differentiate and become active in the taking up and processing of antigens, and their subsequent presentation on the cell surface linked to major histocompatibility (MHC) molecules. Upon appropriate stimulation, DCs undergo further maturation and migrate to secondary lymphoid tissues where they present antigen to T-cells and induce an immune response.
Macrophages
Macrophages are part of the innate immune response. Unlike T and B cells, they do not contain any specific receptors. Macrophages have an important function of homeostasis as they continuously phagocytose self-proteins and cells in their vicinity, during normal tissue repair and aging (e.g. old red blood cells). All of these proteins are degraded and presented on MHC-II. As they are self-proteins so they do not activate T-cells, because, in the absence of infection, macrophages express low levels of MHC-II, and almost no co-stimulator (B7). Further, T cells with high-affinity receptors for self-peptides have been deleted during T cell development in the thymus.
When there is infection, macrophages possess certain types of receptors that recognize differential carbohydrate patterns on foreign cells. They also have receptors for specific bacterial products such as lipopolysaccharide (LPS) (endotoxin). When these molecules bind their bacterial ligands, macrophages become strong antigen-presenting cells because of the up-regulation of MHC-II and B7. They also start to secrete cytokines that aid in their functions (IL-1, 6, 8, 12 and TNF-α). It is at this point that antigen presentation by MHC II will activate Th cells.
B-cells
B-lymphocytes are generated from the common lymphoid progenitors, which are originated from the differentiation of the hematopoietic stem cells. The yolk sac, fetal liver, and the adult liver are the sites in the body where these cells are generated and developed. B-lymphocytes play crucial roles in host defense against infection via a series of highly coordinated processes that include cell homing, antigen recognition, antibody secretion, antigen presentation, and/or cytokine release.
The steps of maturation of B-cells are as follows
In bone marrow in early stages, the B-cell precursors (pre-proB cells) must interact physically with the stromal cells in order for proliferation and maturation to occur. Later stages (late pro-B-cells) merely need the soluble growth factors produced by stromal cells. Stromal cells produce several necessary growth factors and cell-cell adhesion molecules. One key growth factor for B-lymphopoiesis is interleukin-7. The earliest identifiable stage of B-cell differentiation is …… Contents available in the book………… Contents available in the book…… Contents available in the book…… Contents available in the book……
A newly formed B-cell displays IgM on the cell surface. At this stage, the B-cell is still immature and responds to antigen differently from a mature B cell. Immature B-cells can be functionally removed by interaction with self-antigen, either by undergoing programmed cell death (apoptosis) or by anergy, in which the cell is rendered nonresponsive in the presence of the antigen. Thus, similar to T-cells, immature B-lymphocytes undergo a process of “negative selection” to delete cells that are reactive to the “self” antigen. Immature B-cells that are not removed by the processes of negative selection leave the bone marrow and migrate to peripheral or secondary lymphoid tissues such as the spleen and lymph nodes. Here further maturation takes place and the newly formed B-cells express IgD, in addition to IgM, on the cell surface. The mature B-cells are now fully responsive to antigens and interaction with T-cells.
Conclusion
The cellular immune response is the backbone of the overall immune response. It plays a key role in mounting resistance to the infection. It must be remembered hare that the T-cells and B-cells work hand in hand. The immune system as a whole works together to fight infection. In the next article, we shall read about a very important component of the immune system, that is, the complement system.